* Characteristics of living organism
① 생물은 구조적으로 매우 복잡하며 고도로 조직화되어 있다. 생물은 복잡한 내부구조를 가지고 있고, 복잡한 종류의 분자들을 포함하고 있다. 반대로 우리의 환경에 있는 무생물(예컨대, 진흙, 모래, 바위 및 바다물)은 상대적으로 간단한 화학적 화합물로 구성되어 있다.
1. 진정세균(Eubacteria)과 고세균(Archaebacteria)의 차이
2. 메탄 생성 세균[The Methanogens (methane producing bacteria or methanogenic bacteria)]
3. 호염성 세균(The Halophiles)
4. 호열호산성 세균(The Thermoacidophiles)
1. Eubacteria와 Archaebacteria의 차이의 요약 설명
* 세포화학적 차이
- 막 구성지질(인지질) : 리피드(lipid)의 차이
eubacteria : 진핵생물(eukaryote)처럼 글라이세롤(glycerol)간의 탄화수소들이 에스테르 결합을 하고 있다.
archaebacteria : 이소프레노이드 곁사슬('측쇄'라고도 부름)(isoprenoid side chain)이 존재하며, 에테르 결합을 하고 있다.
- 펩티도글리칸(Peptidoglycan)(murein이라고도 함)
archaebacteria의 peptidoglycan : 슈도뮤레인(pseudomurein)(여기서, 'pseudo'란 '허위' 또는 '거짓'을 뜻함)
당유도체(backbone) → eubacteria ; G(N-acetylglucosamine)와
; M (N-acetylmuramic acid)으로 구성되어 있으며, 이 둘사이는 β(1,4) 글리코시드 결합(glycosidic bond)으로 결합되어 있다.
→ archaebacteria ; G(N-acetylglucosamine)와
; T(N-acetyltalosaminuronic acid)로 구성되어 있으며, 이 둘사이는 β(1,3) 결합으로 결합되어 있다.
* 분자 생물학적 차이
-RNA 효소
-tRNA 효소에서의 차이가 있다.
*계통분류학적 차이(Phylogenic Taxonomy)
-16s rRNA(계통분류학적 분석; 언제 분기했는가)
; 핵산을 비교, 분석하는 것이 권장된다.
; DNA가 좋은 재료이지만 거대분자(macromolecule)이므로 rRNA이용한다.
; SJ(Simiarity Coefficient) : 유사도 지수, 범위 : 0.0-1.0(1.0가까울수록 relatedness높다)
;eubacteria사이 SJ = 0.2 or larger
;archaebacteria사이 SJ = slightly less(0.2)
;between eubacteria with archaebacteria SJ = 0.1 or less
⇒0.2보다 값이 낮은 것이 중요, 즉 relatedness가 멀다(없다)
1) 진정세균과 고세균의 차이 정리
① 인지질내의 결합
일반세균 : 지방산과 글리세롤사이에 ester결합
고세균 : ether결합
② R group에 일반세균은 fatty acid가 오는데(single bond) 고세균은 소수성 side chain을 가진 isoprene(systematic name : 2-Methyl-1,3-butadiene)이 온다(double bond; ∵alkene group)
③고세균은 2가지의 인지질 구조가 있다.
1)Glycerol diether
glycerol-ether linkage-isoprenoid chain
-ether linkage-isoprenoid chain → 지질 이중층(lipid bilayer)
2)Diglycerol tetraether
-ether linkage-isoprenoid chain-ether linkage-glycerol
glycerol -ether linkage-isoprenoid chain-ether linkage- → 지질 단일층(lipid monolayer)
2) 고세균의 구성군
고세균은 대사적 또는 생태학적 특성에 근거해서 세 개의 주요군으로 나눌 수 있다.
① 메타노젠;메탄생성균[methanogen(methane producing bacteria)]
: 독특한 에너지 대사의 최종 산물로서 메탄을 생성한다.
② 호염균(halophiles)
서식지 : 높은 염도를 나타내는 환경
③ 호열호산성균(thermoacidophiles)
서식지 : 고온 및 낮은 pH
3) 고세균의 지질
* 고세균에서는 2가지 유형의 지질 구조가 발견된다.
- 글리세롤 디에테르 및 디글리세롤 테트라에테르(glycerol diether and diglycerol tetraether)
* 일반적으로 thermoacidophiles는 주로 dibiphytanyl tetraether를, halophiles는 주로 diphytanyl diether를 가지고 있다. Methanogen에서는 2가지 다른 패턴으로 나누어진다. coccoid cell은 diphytanyl diether만을 가지고 있는 반면, 나머지는 diphytanyl diether와 dibiphytanyl tetraether를 모두 가지고 있다.
2. 메탄생성균[The Methanogens (methane producing bacteria or methanogenic bacteria)]
유기 물질의 부식이 일어나고 있는 대부분의 혐기성 환경에서 생물학적인 메탄 생성은 지구과학적으로 중요한 의미가 있다.
* 환경 : 늪, 호수침전물, 동물의 장내부분, 혐기성 하수 부식물질.
이것은 다른 혐기성 세균에 의해 형성된 발효 산물(CO2, H2, formate, acetate)을 메탄 또는 메탄과 CO2로 변환시키는 매우 특수한 세균군의 활동에 의해 일어난다.
메탄은 물에 잘 녹지 않는 기체상태이기 때문에 혐기성 환경을 벗어나서 혐기성 및 호기성 조건이 접하는 영역에서 사는 호메탄균과 같은 세균군에 의해 호기적으로 산화될 수 있다.
* Mesophiles : 메탄과 같은 단순한 유기화합물(탄소하나로 이루어진 화합물)을 분해하여 에너지 이용하며, 메탄을 섭취(?)하여 이산화탄소로 분해(↑)시키는 세균.
Methanogen은 혐기성 생태계에서 다량의 유기물질이 격리되는 것을 막아주는 역할을 한다.
그대로 두었을 경우, 순환이 안될 탄소를 methanogen이 순환시킨다.
즉, 탄소의 순환에 중요한 역할을 한다.
1) 독특한 보조인자
Methanogen은 다른 세균에서 발견되지 않는 몇가지 보조인자를 가지고 있다.
그들 중 셋[메타노프테린(methanopterin), 메타노푸란(methanofuran) 및 CoM]은 이산화탄소가 메탄으로 환원될 때, C1 unit의 운반체이다; Factor 420은 이 환원과정에서 수소 운반체로서 기능을 하며 Factor 430(an unusual nickel tetrapyrrole)은 이 환원과정에서 마지막 효소인 methyl-CoM reductase의 prosthetic group이다. 일부 methanogen은 생장 인자로서 CoM을 필요로 한다.
#prosthetic group : apoenzyme에 강력히 결합되어 있는 상태의 cofactor or coenzyme을 말함.
2) 에너지 대사
Methanogen이 에너지원으로서 이용하는 화합물은 한정적이다. 일부 포르메이트, 아세테이트, 메탄올, 메틸아민을 에너지원으로 이용하는 균주도 있지만 대부분의 methanogen은 전자공여체로서 H2를 그리고 전자수용체로서 CO2를 에너지원으로 이용한다. ATP합성과 연관된 메탄생성 메카니즘(methanogenesis mechanism)은 알려지지 않았지만, 대부분의 methanogen에는 시(사이)토크롬(cytochrome)과 퀴논(quinone)이 없다. 따라서 통상적인 전자전달은 일어나지 않는다. 그럼에도 불구하고 chemiosmotic membrane potential에 의해 에너지 보존이 중계되는데, 이는 양성자 구배(proton gradient)를 와해시키는 약품을 처리했을 때 ATP농도가 급격히 감소하는 것으로 알 수 있다.
* methanogen에서의 양자구동력(protonmotive force) 형성
- 세포막 밖의 H2가 2H+로 산화되고 분리된 2e-는 막내의 hydrogenase라는 효소에 전달된다.
- 산화형의 F420이 막의 안쪽의 2H+와 결합하여 환원형의 F420이 된다.
- 환원형의 F420은 methanogenesis의 각 부분(필요한 부분)에 2H를 공급한다.
- 막 밖은 [2H+]가 증가하게 되고 막 안쪽은 [2H+]가 감소하게 되어 proton gradient가 형성되고 protonmotive force가 형성된다.
# protonmotive force : an energized state of a membrane created by expulsion of protons usually occurring through action of an electron transport chain.
Protonmotive force는 막에 결합된 ATPase에 의해 ATP로 합성된다. ATPase는 막의 안쪽에 여러개 소단위로 구성된 머리부분과 막의 안과 밖에 결쳐있는 꼬리 부분으로 구성되어 있다. ATPase는 가역반응을 촉매하여 양자구동력을 소비함으로써 일부를 ATP합성에 이용하기도 하고 ATP를 가수분해하여 H+를 밖으로 방출하기도 한다.
* 이 과정에 영향을 주는 chemicals
- inhibitors : 전자전달 방해 ;inhibitors block electron flow.
- uncouplers : 전자전달을 방해하진 않지만 proton gradient를 와해시킴.
(;uncouplers dissipate the proton motive force and hence inhibition of ATP synthesis.)
막에는 ATP → ADP + Pi의 가역반응을 촉매하는 특수한 효소 ATPase가 있기 때문에 세포가 양자 구동력을 ATP합성에 이용할 수 있다. 양자 구동력은 ATPase의 활성을 통해 ATP합성에 이용된다.
# Chemiosmosis : the use of ion gradients, especially proton gradients, across membranes to generate ATP.
3) 황산염에 의한 메탄생성의 저해
* 황산염이 풍부한 환경(바다)에는 황환원세균이 많이 존재하는데, 황산염을 환원시키기 위해 H2를 필요로 하기 때문에, 이러한 환경에서 황환원 세균은 methanogen과 H2를 경쟁적으로 취하려 하며, 이 결과 methanogenesis를 저해하는 요인이 된다.
4H2 + SO42-  H2S + H2O
H2S + H2O
황환원 세균
따라서 바다보다는 담수와 육지에서 methanogenesis가 일어난다.
황환원세균은 전자공여체로 메탄생성균과 똑같은 H2를 이용하기 때문에, 경쟁으로 인해 methanogen에 H2공급이 원활치 않게 된다.
# 최종 전자 수용체에 따른
①(aerobic) respiration(호기적 호흡) : O2
②fermentation(발효) : organic compound(세포내 유기화합물)
③anaerobic respiration(혐기적 호흡) : O2이외의 무기물 ex)SO4(황산염), NO3(질산염)
⇒①과 ③은 최종 전자 수용체(final electron acceptor)가 외부에서 공급되며 ②는 외부에서 공급되지 않는다.
⇒황환원세균(sulfate reducing bacteria)은 혐기적 호흡을 하는 생물체이다.
4) Carbon Assimilation(탄소동화작용)
* 대부분의 methanogen은 유일한 탄소원으로 이산화탄소를 이용한다(독립영양생물: autotrophs).
* 탄소동화회로는 Calvin-Benson cycle과 reductive TCA cycle이 대표적이나 methanogen에서는 발견되지 않으며, 대신 Carbon monoxide pathway가 발견되었다 →이산화탄소 환원.(found in some sulfur-reducing bacteria and some clostridia.)
5) 생태학
* Methanogen은 다량의 유기물이 혐기적 상태에 격리되는 것을 막아주는 역할을 함으로써 탄소의 순환에 중요한 역할을 한다.
* Hydrogenase : Km값이 매우 낮다.
Km은 친화도(affinity)에 관한 것으로 속도가 증가할 수록 Km값은 줄어들고 속도가 줄어들수록 Km은 증가한다. 효소에 따라 기질과의 친화도가 높고 낮음을 나타낸다. 친화도가 높을수록 Km값은 낮아진다.
* Methanogen의 hydrogenase는 Km값이 낮다, 즉 affinity가 높다. 따라서 주위의 수소분압은 소모되어 낮은 상태를 유지한다.
* 혐기성세균의 H2이용
NADH + H+  NAD+ + H2
NAD+ + H2
혐기성세균이 메탄생성균과 함께 존재하면 메탄생성세균이 H2를 소모해버리기 때문에 화학반응의 평형이 오른쪽으로 치우친다.
NADH + H+  NAD+ + H2
NAD+ + H2
NADH보다 NAD+가 더 많이 만들어지게 되어 산화반응이 더욱 진행되므로 정상때보다 더 산화된 최종 산물이 생성된다.
높은 수율의 ATP와 biomass가 만들어진다. 이와 같은 현상을 interspecies hydrogen transfer(종간수소전달)라고 한다.
<중요> Interspecies hydrogen transfer
Methanogen의 hydrogenase의 Km값은 활발한 methanogenesis가 일어나기에 충분할 만큼 낮은 H2분압을 유지시켜주며 이러한 낮은 H2분압은 혐기성세균이 NADH를 다시 산화시키도록 한다. 이러한 hydrogenase는 혐기성세균에 널리 분포되어있다. 혐기성세균을 순수배양시키면 화학평형이 왼쪽으로 치우쳐 있기 때문에 NADH의 재산화가 이루어지지 않는다. 그러나 메탄생성균과 함께 배양시키면 메탄생성균의 methanogenesis로 인해 연속적으로 H2가 소모되어 낮은 H2분압이 만들어지며 혐기성세균은 NADH를 산화시켜 NAD+를 많이 만들게 되어 산화반응이 더욱 진행되므로 정상때보다 더 산화된 end product가 생산되고 높은 수율의 ATP와 biomass가 증가하게 되어 결국 cell은 활발하게 증식하게 된다. 이러한 현상을 interspecies hydrogen transfer라고 한다.
3. 호염성 세균
1) 호염성 세균에서의 광인산화
대부분의 halophile은 절대적으로 호흡물질대사를 하지만 그 중 일부는 특이한 photophosphorylation의 형태(방식)에 의해 ATP를 합성할 수 있다. Halobacteria의 경쟁하는 종들이 산소가 제한된 상태에 놓이게 되면 화학적으로 변화된(수정된) 세포막을 합성한다. 호기적으로 성장한 세포에서 막은 많은 캐로티노이드(carotenoid)성분으로 인해 적색을 띤다. 하지만, 혐기적환경에선 생장을 멈추며 새로운 구성요소인 자색막(purple membrane)의 합성이 유도된다.
* Halobacterium속에 속하는 세균중 일부는 특이한 광인산화반응을 통해 ATP를 합성할 수 있다. Halobacterium은 호기적 환경에선 호기적호흡에 의해 아미노산을 이용하여 ATP합성한다. 혐기적 환경에서 살게되면 성장을 멈추면서 carotenoid에 의해 red를 띠던 membrane이 purple을 띠게 된다.
Purple membrane은 25%의 지질과 한 가지의 단백질(bacteriorhodopsin)으로 구성되어 있다.
박테리오로돕신(Bacteriorhodopsin)은 혐기적 환경에서 새롭게 나타난 단백질로서, 이 단백질 의해 혐기적 환경의 halophiles은 purple을 띠게 되며 빛을 이용하여 ATP를 생성한다. Bacteriorhodopsin의 크로모포어[chromophore(발색단;빛에너지를 받아들이는 부위)]는 bacteriorhodopsin의 리신(lysine)잔기의 epsilon amino group(즉, 5번째 아미노잔기를 말함)에 쉬프 베이스(Schiff base)를 통해서 연결되어 있는 캐로티노이드 레티널(carotenoid retinal)이다.
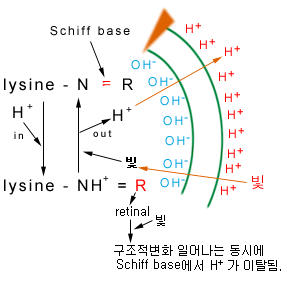
Bacteriorhodopsin은 양성자 펌프(proton pump)로서 역할을 하며 빛 에너지를 proton motive force로 전환시켜 세포의 ATP level을 유지시킬 수 있다.
그러나, 성장은 못한다.
∵β-carotenoid에서 retinal이 만들어지는데, 그 과정에서 반드시 O2를 요구한다. 성장(=division)을 위해선 모(母)세포에서 retinal이 2배가 되야 한다. 하지만 더 이상 O2가 없기 때문에 retinal을 만들지 못하여 분열할 수가 없다.
최초의 retinal을 만들기 위해 수중에 남아있는 O2, 세포내의 O2를 모두 소비한다.
Halobacterium은 혐기적 환경에서 빛을 이용하여 ATP를 생성할 수는 있지만 이러한 환경에서 성장할 수는 없다. Photophosphorylation은 비성장 세포(nongrowing cell)의 생존력의 연장에만 사용된다. Halobacterium이 빛이 있는 혐기적인 환경에서 성장할 수 없다는 것은 β-carotene으로부터 retinal을 형성하기 위한 O2분자의 요구도를 시사한다.
따라서 광종속영양적 성장(photoheterotrophic growth)는 O2분압이 bacteriorhodopsin의 합성을 유도할 만큼 낮지만 retinal을 생합성할 정도로 높을 경우에만 일어난다.
4. The Thermoacidophiles
* The thermoacidophiles are defined by their ability to grow at high temperature and low pH.
* Subgroups - Sulfolobus
- Thermoplasma
- Thermoproteus
1) Sulfolobus
* facultative chemoautotroph
* 서식지 : 고온 산성 온천 및 토양
* 최적 생장 온도 : 63-80℃
pH : 2.0
* 생장 온도 범위 : 55-85℃
pH : 1.0-5.9
* 황원소를 환원시킴.
2) Thermoplasma
* facultative anaerobe
* 탄소원 및 에너지원으로서 소수의 단당류 및 이당류를 이용
* 잔여 석탄 및 상당량의 황철광(FeS)을 함유한 탄광의 쓰레기 더미에서만 발견됨
⇒oxidation of the FeS by chemoautotrophic bacteria acidifies and heats the pile, creating a favorable growth environment for Thermoplasma(pH 0.5 to 4.5 ; temperature 37 to 65℃).
3) The Thermoproteus Group
* 최종 전자 수용체로서 황원소를 사용하는 물질대사를 원칙적으로 또는 유일한 호흡 대사로 이용함.
* 이들의 물질 대사 방법은 호흡 기질로서 다양한 당류, 알콜, 유기산(fromate 포함) 및 CO를 사용하는 호흡이다.
* 온도 : 78 내지 95℃.
* pH : 2.5 내지 6.
'학술관련 > bacteria' 카테고리의 다른 글
| Spore Forming Bacteria (1) | 2008.12.23 |
|---|---|
| BL21-DE3, BL21-DE3-RIL (1) | 2008.07.02 |
| E.coli BL21 plysS와 E.coli BL21 (0) | 2008.03.25 |
| BL21 Competent Cells for Expressing Sequenced Constructs (0) | 2007.09.06 |
| 대장균군 (Coliform Group) (1) | 2007.09.06 |